Автор — Маргарита Шувалова, сотрудник Федерального центра мозга и нейротехнологий, аспирант и младший научный сотрудник Института биоорганической химии Российской академии наук; Анастасия Фокина, студентка 4 курса ФББ МГУ, преподаватель биологии в АПО.
Строение ДНК
Чтобы понять, как протекает репликация, нужно знать строение той молекулы, которая будет реплицироваться. У всех без исключения клеточных форм жизни генетическая информация хранится в форме двойной спирали ДНК (дезоксирибонуклеиновой кислоты). Однако существуют ещё и вирусы — неклеточные формы жизни, некоторые из которых хранят генетический материал в виде РНК (рибонуклеиновой кислоты). О репликации вирусных геномов мы поговорим в других статьях, а пока что сосредоточимся на организмах, геном которых хранится в виде молекулы ДНК.
История открытия
ДНК была открыта Фредериком Мишером в 1869 году, и на протяжении почти ста лет её структура оставалась для ученых загадкой. И только в 1953 году Френсису Крику и Джеймсу Уотсону (рис. 1) удалось с использованием рентгеноструктурных данных, полученных Розалиндой Франклин (рис. 2), установить, что ДНК существует в виде двойной спирали (рис. 3). За это открытие Уотсон и Крик в 1962 году удостоились Нобелевской премии, а Франклин, к огромному сожалению, скончалась от болезни четырьмя годами ранее и не была отмечена данной наградой, потому что посмертно ее не вручают. Позже Джеймс Уотсон написал автобиографическую книгу «Двойная спираль», где рассказал об истории их открытия.


Рис. 3. Изображение ДНК, полученное с помощью рентгеноструктурного анализа, по которому была установлена структура этой макромолекулы. Источник.
Строение нуклеотидов и связи между ними
ДНК (дезоксирибонуклеиновая кислота) — линейный полимер, состоящий из мономеров — нуклеотидов. Нуклеотиды ДНК называются общим словом дезоксирибонуклеотиды (нуклеотиды, входящие в состав РНК, называются просто рибонуклеотиды). Каждый дезоксирибонуклеотид состоит из трех компонентов: азотистое основание, пятиуглеродный сахар (дезоксирибоза) и остаток фосфорной кислоты (рис. 4).

Азотистые основания в дезоксирибонуклеотидах бывают четырех видов: аденин, тимин, гуанин и цитозин. Два из них — аденин и гуанин — с химической точки зрения являются производными вещества пурина, поэтому носят название пуриновые основания. Два оставшихся — тимин и цитозин — производные пиримидина, поэтому они называются пиримидиновыми основаниями.
В ДНК никогда в норме не встречается пиримидиновое основание урацил. Однако он может спонтанно образоваться из цитозина в результате химической модификации, тогда клетка распознает его как мутацию и постарается его вырезать. Именно поэтому в ДНК находится не урацил, а тимин — тимин имеет дополнительную метильную группу, благодаря которой клетка точно знает, что это правильное азотистое основание, а не результат случайного химического превращение цитозина.
Второй компонент нуклеотида — это пятиуглеродный сахар. В дезоксирибонуклеотидах это дезоксирибоза. От рибозы она отличается тем, что вместо гидроксила у 2 атома углерода она содержит водород. Отсутствие гидроксильной группы в дезоксирибозе делает ДНК более устойчивой к распаду, что, конечно же, очень важно для молекулы-хранительницы генетической информации. Возможно, дезоксирибоза появилась в ДНК вследствие естественного отбора, направленного на повышение устойчивости данной молекулы.
Азотистое основание и дезоксирибоза связаны между собой N-гликозидной связью (то есть связь идет через азот). Фрагмент нуклеотида, представляющий из себя азотистое основание, связанное с сахаром, называется нуклеозид.
Для того, чтобы не путать атомы кольца основания и кольца дезоксирибозы, при нумерации к атомам дезоксирибозы добавляют штрих: 1’, 2’ и так далее. Атомы основания нумеруют без штриха.
Третий компонент нуклеотида — это остаток фосфорной кислоты. Если нуклеотид уже встроен в ДНК, то он имеет один такой остаток, а если же он находится в свободном виде, то у него их три, соединенных фосфоангидридными связями. Эти связи макроэргические, т.е. при их разрушении выделяется энергия, которая может быть затрачена на какие-то процессы. В данном случае разрушается связь между первым и вторым остатками фосфорной кислоты, выделяется пирофосфат, а энергия уходит на присоединение нуклеотида к растущей цепи ДНК. Получается, что энергия, необходимая на построение полимера, заключена в самих мономерах (как если бы в кирпичах содержалась энергия для постройки стены). К глубокому биологическому смыслу, заложенному в этом явлении, мы ещё вернемся.

Встраивание нуклеотида в растущую цепь ДНК происходит следующим образом: атом кислорода у 3’ атома дезоксирибозы предыдущего нуклеотида осуществляет нуклеофильную атаку на фосфат в присоединяемом нуклеотиде. В результате выделяется пирофосфат, который быстро разлагается до ортофосфата, что сдвигает направление реакции в сторону полимеризации (рис. 5).
В цепи ДНК нуклеотиды соединены между собой через остаток фосфорной кислоты. У первого нуклеотида цепи свободен остаток фосфорной кислоты у 5’ углерода, так как он ни с кем не образует связь через него, а у самого последнего — свободен 3’ гидроксил. Соответственно, передний конец цепи получил название «5’-конец» («пять штрих»), а задний конец — «3’-конец». Таким образом, цепь ДНК имеет «начало» и «конец».
Двойная спираль ДНК состоит из двух антипараллельных цепей: там, где у одной цепи находится 5’ конец, у второй — 3’-конец (рис. 6).

Цепи в двойной спирали удерживаются благодаря водородным связям, возникающим между комплементарными основаниями двух цепей, а также благодаря стекинг-взаимодействиям соседних нуклеотидов одной цепи. Между аденином и тимином образуется две водородные связи, между гуанином и цитозином — три. Для того, чтобы разрушить пару Г-Ц, нужно больше энергии, т.к. в ней на одну водородную связь больше, чем в А-Т. Комплементарные пары А-Т и Г-Ц называют Уотсон-Криковскими парами, благодаря их формированию возможна репликация ДНК (рис. 7).

Строение двойной спирали
Двойная спираль может существовать в нескольких формах, наиболее важная и широко встречающаяся — это В-форма. На один виток спирали приходятся 10 пар нуклеотидов, виток имеет длину 3,4 нм, спираль правозакрученная. Известно ещё довольно много форм ДНК, которые называются буквами английского алфавита, формы А и Z показаны на рисунке 8. Подробнее о разнообразии форм ДНК можно прочитать в этой статье.

Механизм репликации
Мы кратко познакомились со структурой молекулы ДНК, а теперь перейдем непосредственно к её репликации.
Репликация ДНК протекает по полуконсервативному механизму. Это означает, что одна из цепей синтезируется заново, а другая остается от исходной молекулы. Про эксперименты, которые это доказали, а также про другие выдвинутые гипотезы репликации ДНК можно прочитать тут.
Полуконсервативный механизм репликации логически вытекает из строения ДНК, а точнее — из комплементарности оснований. Вторую цепь ДНК можно однозначно построить на информации, заключенной в первой цепи, используя правило комплементарности. Обратите внимание, что вторая цепь не идентична первой, она сопоставлена ей по определенному правилу, закодирована в ней. Интересно, что все матричные синтезы, существующие в природе (репликация, прямая и обратная транскрипция, трансляция), всегда используют какое-то правило сопоставления и никогда не делают просто вторую идентичную цепь или молекулу на основе первой.
Наращивание цепочки нуклеотидов происходит в направлении 5’ -> 3’. Это означает, что новые нуклеотиды своими фосфатами присоединяются к 3’ гидроксилам предыдущего нуклеотида и никогда (или почти никогда?*) наоборот.
*нам удалось найти всего одно упоминание роста нуклеотидной цепи в направлении 3’ ->5’. Она происходит при созревании некоторых тРНК, то есть речь идет о рибонуклеиновой кислоте. Таким способом происходит присоединение всего нескольких нуклеотидов. Особо заинтересованным читателям можем предложить прочитать статью на английском языке об изучении этого феномена.
Это связано с тем, что энергия, требующаяся для наращивания цепи, содержится в самих нуклеотидах, в их фосфоангидридных связях. Дело в том, что такая связь довольно легко гидролизуется, поэтому есть всегда вероятность ее «поломки». Если эта «поломка» происходит в нуклеотиде, то ничего драматичного не случается — нуклеотидов в клетке множество, клетка просто возьмет новый и продолжит репликацию. А вот если бы направление репликации было бы от 3’ к 5’ концу, это значило бы, что остатки фосфорной кислоты находились бы на растущей молекуле ДНК. Каждая их «поломка» носила бы критический характер, ведь ДНК в клетке содержится в единственном экземпляре (в ди- и полиплоидных клетках в нескольких экземплярах, но каждый из них уникален и терять его не хочется)! Видимо, это ограничение носит очень жесткий характер, ведь даже в тех случаях, когда, казалось бы, удобнее было бы синтезировать в направлении 3’ -> 5’, клетка всё равно этого не делает.
Ферменты, участвующие в репликации
Репликация ДНК не происходит самопроизвольно, это сложный процесс, осуществляемый множеством ферментов.
Репликация начинается с инициации, в которой участвуют хеликаза, топоизомераза и белки, связывающие одноцепочечную ДНК (SSB-белки).
Чтобы построить комплементарную цепь, сначала нужно разорвать водородные связи в исходной двойной спирали. Разрыв водородных связей между цепями ДНК — «плавление» — самопроизвольно протекает только при температуре, близкой к точке кипения воды. В клетке, разумеется, такие температуры не достигаются, а за разрушение водородных связей отвечает фермент хеликаза («хеликс» — спираль, фермент, разрушающий спираль). Хеликаза представляет из себя кольцо, которое движется по одной из цепей ДНК с использованием энергии АТФ. Это движение разрывает водородные связи и делает основания доступными для образования новых комплементарных пар.
Для того, чтобы две цепи не «слиплись» обратно, а цепи ДНК не образовали шпильки сами с собой, существуют так называемые SSB-белки (от английского single strand binding, «связывающие одну цепь»). Как следует из названия, они связываются с отдельными цепями ДНК и не дают им заново образовать двойную спираль.
Расхождение двойной цепи в одном месте неизбежно вызывает её деформацию в другом месте. Это хорошо иллюстрирует канат: если вы попытаетесь разделить волокна каната в одном месте, то в другом месте происходит его избыточное скручивание. Чтобы этой деформации не происходило, работают ферменты топоизомеразы. Топоизомераза I делает одноцепочечный разрыв, а топоизомераза II — двухцепочечный. «Прокручивая» молекулу ДНК, они снимают с нее избыточную спирализацию. Топоизомераза II также может разделять «запутавшиеся» ДНК, чтобы они не порвались при делении клетки.
Хеликаза, топоизомераза и SSB-белки, взаимодействующие с ДНК, изображены на рис. 9.

ДНК-полимераза
Основной фермент, который участвует в элонгации репликации, — это ДНК-полимераза. Именно она достраивает дочернюю цепь на материнской по правилу комплементарности. Сама по себе ДНК-полимераза может синтезировать только короткие фрагменты ДНК и потом отваливается от реплицируемой цепи. Ее активность повышает скользящий зажим — белковое кольцо, которое фиксирует ДНК-полимеразу на нити ДНК и повышает её процессивность (рис. 10).

ДНК-полимераз существует великое множество. На основании своей структуры они делятся на семь семейств — A, B, C, D, X, Y, и RT. Некоторые из них не участвуют в репликации, а нужны только для процессов восстановления ДНК — репарации или для обратной транскрипции. Некоторые из них еще очень плохо изучены и их роль в клетке не до конца понятна. Полимеразы, которые участвуют в удвоении ДНК, называют репликативными, а те, которые задействованы в исправлении повреждений ДНК, — репаративными, однако это деление условно. Еще большую путаницу в разнообразие ДНК-полимераз вносит то, что у бактерий, архей и эукариот они разные. Чтобы лучше разобраться со всем многообразием ДНК-полимераз, рекомендуем ознакомиться вот с этой книгой. Мы сосредоточимся на описании репликативных ДНК-полимераз бактерий и эукариот.
У бактерий известно по крайней мере 5 ДНК-полимераз. Они нумеруются римскими цифрами от I до V. Основной репликативный фермент — это ДНК-полимераза III (относится к семейству С). Она состоит из нескольких отдельных субъединиц — α, β, γ, δ, δ’, ε, θ, τ, χ, ψ. α-субъединица имеет полимеразную активность, ε-субъединица отвечает за корректировку неправильно включенного нуклеотида (3’->5’ экзонуклеазная активность), а β-субъединица является «скользящим зажимом». Остальные субъединицы нужны для правильного взаимодействия этих субъединиц и для повышения их активности.
Структурная модель ДНК-полимеразы, напоминающая по форме ладонь, представлена на рис. 11.

Репликативная ДНК-полимераза не может сесть на конец ДНК и начать синтез с нуля, для начала работы ей обязательно требуется затравка — праймер. Праймер представляет собой короткий фрагмент из РНК, который синтезируется ферментом праймазой. Праймер потом вырезается РНКазой Н, и на его месте достраивается комплементарная цепь уже из ДНК (рис. 12). Та же праймаза синтезирует фрагменты Оказаки на отстающей цепи, что будет обсуждаться далее.

Почему для репликативной ДНК-полимеразы необходим праймер? Неужели в процессе эволюции нельзя было «изобрести» такую ДНК-полимеразу, которая была бы «самодостаточной»? Ответ кроется в самокорректирующей способности ДНК-полимеразы. Она обязательно должна быть связана с предыдущим нуклеотидом, чтобы проверить его на правильность и в случае необходимости вырезать его, заменив на нужный. Таким образом, клетка производит всю эту сложную систему с праймерами для того, чтобы повысить точность самой репликации. Сам праймер, кстати, синтезируется ферментом без самокорректирующей активности, поэтому в нем относительно много ошибок. Но это и неважно, ведь далее он вырезается. Выбор именно РНК в качестве праймера облегчает процесс его удаления: РНК в составе ДНК автоматически распознается клеткой как чужеродный компонент и расщепляется.
Для удаления праймера бактерии используют ДНК-полимеразу I. Она обладает способностью вырезать праймер, достраивая на его месте комплементарную цепь ДНК.
У эукариот все еще более запутанно. Для них известно 15 (!) различных видов ДНК-полимераз, которые обозначаются греческими буквами, начиная с альфы. Некоторые из них ответственны за репарацию, а функции нескольких вообще остаются загадкой. Мы поговорим о двух самых важных ДНК-полимеразах — альфе и дельте.
ДНК-полимераза дельта (δ) — основная полимераза репликации, она делает практически все работу по наращиванию дочерних цепей ДНК. Она может исправлять сама себя, т.е. обладает 3’ ->5’ экзонуклеазной активностью. ДНК-полимераза δ состоит из многих субъединиц: каталитических, регуляторных, а также тех, чьи функции еще не до конца ясны. Для ДНК-полимеразы δ, как и для прокариотических полимераз, необходим скользящий зажим — кольцо, которое «толкает» фермент вперед и не дает ему раньше времени отсоединиться от ДНК.
ДНК-полимераза альфа (α) имеет две активности — ДНК-полимеразную и РНК-полимеразную. Она синтезирует сначала РНК-праймеры, а потом присоединят к ним несколько дезоксирибонуклеотидов. ДНК-полимераза α не имеет самокорректирующей активности, поэтому и праймер, и тот участок ДНК, который она синтезировала, получаются не очень точными и впоследствии вырезаются, а затем застраиваются ДНК-полимеразой дельта. Таким образом, ДНК-полимераза α выполняет роль праймазы.
Все разнообразие ДНК-полимераз эукариот и прокариот суммировано в таблице 1.
| Семейство | ДНК-полимеразы | Функции |
| А | ДНК-полимеразы I бактерий, ДНК-полимераза фага Т7 | Репликация, репарация |
| B | ДНК-полимераза II E.coli, ДНК-полимеразы фагов Т4 и RB69, некоторые ДНК-полимеразы эукариот, ДНК-полимеразы В архей | Репликация, репарация |
| C | ДНК-полимеразы III бактерий | Репликация, репарация |
| D | ДНК-полимеразы D эуриархеот | Репликация, репарация |
| Y | ДНК-полимеразы IV, V E.coli. ДНК-полимеразы I и REV1 эукариот | Синтез через повреждения |
| X | Некоторые ДНК-полимеразы эукариот | Репликация, репарация |
Инициация репликации, образование репликативной вилки
У бактерий на кольцевой хромосоме обычно есть всего одна точка начала репликации — она называется OriC (от слова origin — начало). На самом деле это не точка, а протяженная область из нескольких сот пар нуклеотидов (у кишечной палочки, например, из 245 пар). Эта последовательность богата парами А-Т, поэтому в этом месте двойная спираль относительно легко расходится (помним, что между А и Т образуется всего две водородные связи).
Белок DnaA играет ключевую роль в инициации репликации у многих бактерий. Он последовательно выполняет 3 главные функции: 1) узнает область начала репликации oriC; 2) способствует «расплетанию» легкоплавких АТ-богатых участков ДНК oriC; 3) привлекает на расплетенные участки oriC хеликазы. Активация хеликаз приводит к «расплетанию» двойной цепи и образованию репликативного «глаза». В «уголках» этого глаза находятся две репликативные вилки, которые движутся в противоположные стороны. На расплетенные нити ДНК садится праймаза, катализирующая образование РНК-праймера, а затем и ДНК-полимераза III вместе со своим скользящим зажимом. SSB-белки связываются с одноцепочечными ДНК, стабилизируют их и не дают им соединиться обратно или образовать шпильки. Таким образом, инициация репликации ДНК на oriC завершается образованием двух движущихся в противоположных направлениях репликативных вилок.
На рис. 13 изображена схема репликации в клетках прокариот с одним ориджином.

У эукариот, в отличие от бактерий, точек начала репликации множество, иначе бы весь геном реплицировался очень долго (например, у дрожжей на одну хромосому может приходиться до 400 точек начала репликации). С ними связывается белковый комплекс ORC — origin recognition complex, комплекс, узнающий начало репликации. На ORC присоединяется хеликаза, однако дальнейшее функционирование репликативного комплекса возможно, только если клетка получит на это разрешение от специальных белков, регулирующих клеточных цикл. Репликация ДНК должна происходить только в S-фазу клеточного цикла, поэтому репликационная машина терпеливо ждет, когда эта фаза наступит. Если клетка успешно вступила в S-фазу, то хеликазы активируются и начинают расплетать ДНК. Далее происходит синтез праймеров и присоединение ДНК-полимеразы.
Репликативные вилки движутся до тех пор, пока не столкнутся лоб в лоб со встречной вилкой или не дойдут до конца хромосомы.
На рис. 14 изображена схема репликативной вилки ДНК.
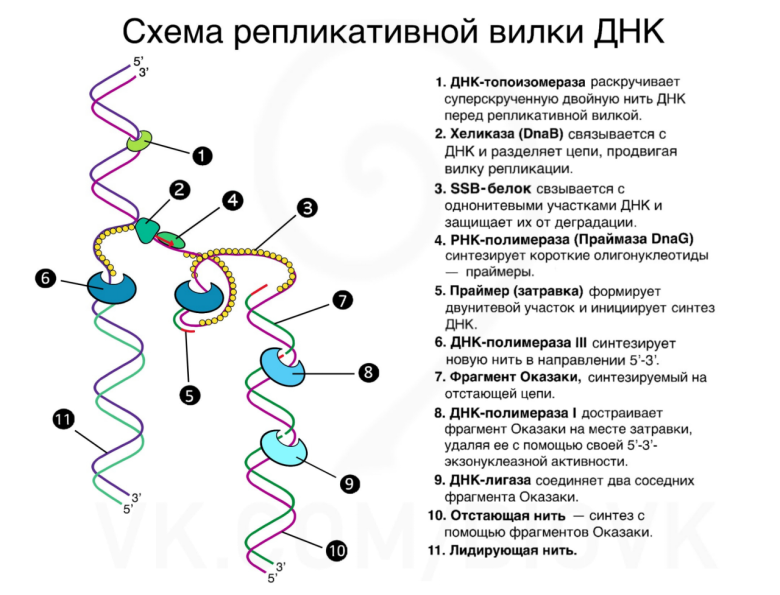
Элонгация репликации. Фрагменты Оказаки
Новые цепи ДНК синтезируются антипараллельно материнским. На одной из материнских цепей синтез идет в направлении 3’ -> 5’ относительно репликативной вилки, следовательно, дочерняя цепь синтезируется на ней непрерывно в направлении 5’ -> 3’. Такая материнская цепь, на которой синтез идет непрерывно, называется лидирующей. Вторая материнская цепь (ее называют отстающей) располагается в направлении 5’ -> 3’ по отношению к репликативной вилке. Первоначально полагали, что комплементарная цепь на ней достраивается непрерывно в направлении 3’ ->5’, но в дальнейшем выяснили, что этого не происходит по причинам, описанным выше. Вместо этого комплементарная цепь достраивается праймазой 8 «обратными стежками» — фрагментами Оказаки (они названы так в честь японской супружеской пары, которая их открыла, — Цунеко и Редзи Оказаки) (рис. 15).

Когда репликативная вилка продвигается, на отстающей цепи открываются новые нуклеотиды с 5’ конца. В направлении, противоположном движению вилки, достраивается сначала праймер, а потом и участок ДНК — фрагмент Оказаки. Далее фрагменты Оказаки сшиваются ферментом лигазой, перед этим та же РНКаза Н расщепляет РНК-праймер, а ДНК-полимераза I достраивает ДНК на месте удаленного праймера. ДНК-лигазы катализируют образование фосфодиэфирной связи в однонитевом разрыве ДНК между соседними нуклеотидами в разорванной нити. ДНК-лигазы участвуют в воссоединении фрагментов в одну непрерывную цепь. Таким образом, на отстающей цепи репликация происходит фрагментами и в направлении, противоположном движению репликативной вилки. Вся эта сложная машинерия была создана лишь с одной целью — обойти запрет на репликацию в направлении 3’ ->5’. Вот насколько жесткое это правило!
Терминация репликации
У эукариот нет специфического сайта для терминации репликации, она происходит при встрече двух разнонаправленных репликативных вилок. В прокариотических хромосомах есть специальные последовательности-терминаторы, которые останавливают синтез ДНК. Эти последовательности позволяют двум репликативным вилкам проходить в одном направлении, но не позволяют проходить в другом.
Из-за того, что ДНК-полимераза не может начать синтез цепи без затравки, с каждым циклом деления происходит недорепликация теломер — концевых участков хромосом. Т.е. с каждым клеточным делением наши хромосомы понемногу укорачиваются. Теоретически это могло бы привести к потере генов, но на концах хромосом расположены длинные некодирующие последовательности, укорочение которых не приводит к потере важной информации. Более подробно о теломерах и о том, как клетка может их наращивать, можно почитать здесь.
Заключение
Итак, давайте попробуем кратко подытожить, что происходит в процессе репликации ДНК.
Репликация инициируется в точке начала репликации, представляющей собой особую последовательность нуклеотидов. С этой последовательностью связываются специальные белки (DnaA — у бактерий, и комплекс ORC у — эукариот) и обеспечивают присоединение хеликаз. Хеликазы расплетают двойную спираль, разрывая комплементарные связи между нуклеотидами. Расплетенную ДНК стабилизируют SSB-белки. Область расхождения цепей двойной спирали называется репликативным глазом. В углах этого глаза находятся репликативные вилки, которые движутся в противоположные стороны. Для начала репликации ферменту ДНК-полимеразе необходима затравка — РНК-праймер. У бактерий он синтезируется праймазой, а у эукариотов — ДНК-полимеразой альфа. Главная репликативная полимераза у бактерий — это ДНК-полимераза III, состоящая из многих субъединиц. ДНК-полимераза I вырезает праймеры и достраивает на их месте цепь ДНК. У эукариот главная репликативный фермент — ДНК-полимераза дельта. ДНК-полимеразы могут исправлять собственные ошибки, вырезая последний неправильно спаренный нуклеотид — 3’ -> 5’ экзонуклеазная активность.
Цепь ДНК, которая ориентирована относительно движения вилки в направление 3’ ->5’, называется лидирующей, синтез комплементарной цепи на ней идет непрерывно. Цепь, ориентированная в направлении 5’ -> 3’, называется отстающей, синтез комплементарной цепи на ней идет фрагментами Оказаки в направлении, противоположном движению вилки. Фрагменты Оказаки сшиваются между собой ферментом лигазой. Впереди репликативной вилки возникает суперскручивание двойной спирали, оно удаляется ферментами топоизомеразами. Репликативный комплекс разрушается, если сталкивается с другим комплексом, идущим в противоположную сторону, или доходит до конца хромосомы.
Источники:
- Альбертс Б. Брей Д., Льюис Дж., Рэфф М., Роберте К. Уотсон Дж. «Молекулярная биология клетки» В 3-х т 2-е изд., Пер. с англ. -М.: Мир, L 994. -504 е., ил. ISBN: 5-03-001985-5.
- Science History Institute: «Francis Crick, Rosalind Franklin, James Watson, and Maurice Wilkins».
- El ADN (I): la molécula, por Rocío Bautista, 26 августа 2019, Biotecnología, Genética.
- КНИТУ (КХТИ): «Нуклеиновые кислоты».
- Полина Лосева, «Как все закручено», N+1.
- Литвинович Татьяна Викторовна, «Репликация ДНК. Транскрипция. Трансляция».
- William A Beard,Samuel H Wilson, Structural Insights into the Origins of DNA Polymerase Fidelity, Мay 2003.
- Molecular Biology. Genes to proteins. «DNA replication», Jones and Bartlett Publishers.
- Материалы группы «Биология» «ВКонтакте», «Схема репликативной вилки ДНК».
- Материалы платформы Khan Academy, «Молекулярный механизм репликации ДНК».
- М. П. Рубцова, Д. П. Василькова и др. «Функции теломеразы: удлинение теломер и не только». 2012 г. УДК 577.57.017.6.
Рекомендуемые материалы:
- Альбертс Б. Брей Д., Льюис Дж., Рэфф М., Роберте К. Уотсон Дж. «Молекулярная биология клетки» В 3-х т 2-е изд., Пер. с англ. -М.: Мир, L 994. -504 е., ил. ISBN: 5-03-001985-5.2.
- М. П. Рубцова, Д. П. Василькова и др. «Функции теломеразы: удлинение теломер
и не только». 2012 г. УДК 577.57.017.6.3. - Полина Лосева, «Как все закручено», N+1.4.
- .Jackman JE, Gott JM, Gray MW. Doing it in reverse: 3′-to-5′ polymerization by the Thg1 superfamily. RNA. 2012 May;18(5.:886-99. doi: 10.1261/rna.032300.112.
- Калинин В.П., «Репликация генома».